Eocen
| Var [ 1 ] | Periode | Epoke | millioner år |
|---|---|---|---|
| Kenozoikum | Kvartær | Holocen | 0,01 (~10 000 f.Kr.) |
| Pleistocen | 2,59 | ||
| Neogen | Pliocen | 5,33 | |
| miocen | 23.03 | ||
| Paleogen | Oligocen | 33,9 | |
| Eocen | 56,0 | ||
| paleocen | 66,0 |
Eocen er en epoke og serie på den geologiske tidsskalaen som tilhører den paleogene perioden og systemet ; innenfor dette følger eocen paleocen og går foran oligocen . Det startet for rundt 56 millioner år siden og sluttet for rundt 34 millioner år siden. [ 2 ] [ 3 ] [ 4 ] [ 5 ]
I løpet av denne tiden begynte noen av de mest betydningsfulle fjellkjedene i verden i dag å dannes, for eksempel Alpene og Himalaya , og flere viktige klimatiske endringer skjedde: det termiske maksimumet av Paleocen-Eocene , som økte temperaturen på planeten og avgrenser begynnelsen på denne geologiske epoken; og Azolla-arrangementet , en global avkjøling som ville gi plass til de første istidene . Grande Coupure -masseutryddelsen markerer slutten på eocen .
Fugler dominerte over andre vesener, og de første hvalene begynte sin utvikling. Videre dateres den største slangearten som har eksistert til eocen, og en stor utvidelse og diversifisering av maur skjedde . Antarktis begynte epoken omgitt av tropiske skoger , og endte den med utseendet til de første polare iskappene. Det er mange paleontologiske steder i forskjellige deler av verden som bekrefter disse fakta, for eksempel fossilområdet Messel i Tyskland , eller Green River-formasjonen i Nord-Amerika .
Navnet Eocene, definert av briten Charles Lyell , kommer fra de greske ordene eos (ἠώς, 'daggry') og kainos (καινός, 'ny'), og refererer til utseendet til de moderne ordener av pattedyr i løpet av denne tiden.
Underavdelinger
Eocen er vanligvis uformelt delt inn i nedre eocen ( ypresisk ) , mellomeocen ( lutetisk og bartonsk ) og øvre eocen ( priabonsk ). Det er sjeldnere å dele det inn i bare underordnet og overordnet. I dette tilfellet ville lutetianeren bli en del av nedre eocen, mens bartonianeren ville bli en del av øvre eocen.
| var Erathema |
Systemperiode _ |
Periodeserien _ |
Gulv alder |
Start, om millioner av år | |
|---|---|---|---|---|---|
| Kenozoikum [ 6 ] | Kvartær [ 6 ] | 2.588 | |||
| Neogen | 23.03 | ||||
| Paleogen |
Oligocen | Chattian | 28.1 | ||
| Rupelian | 33,9 | ||||
| Eocen | Priabonian | 37,71 | |||
| Bartonsk | 41,3 | ||||
| Lutetian | 47,8 | ||||
| ypressense | 56,0 | ||||
| paleocen | Thanetian | 59,2 | |||
| Sjællandsk | ~61,6 | ||||
| dansk | 66,0 | ||||
- Ypresian – begynte for 55,8 ± 0,2 millioner år siden, sammenfallende med begynnelsen av det paleocene-eocene termiske maksimumet , en periode med rask og intens global oppvarming som førtetil at mange bentiske foraminifer døde ut , og som er assosiert med en episode med stor utvikling av pattedyr . I den stratigrafiske serien er begynnelsen preget av en variasjon av 13 C isotopen , siden nivåene av CO 2 økte og forholdet mellom 13 C isotopen i forhold til 12 C redusert. Slutten av denne tidsalderen er preget av en stor utvikling av planktoniske foraminiferer og av utseendet til slekten Hantkenina . Den endte for 48,6 ± 0,2 millioner år siden, og skylder navnet sitt til byen Ypres i Belgia . [ 4 ]
- Lutetian – begynte for 48,6 ± 0,2 millioner år siden. Dette er en tidsalder med rikelig med marine virvelløse dyr ( bløtdyr , koraller , kråkeboller ) og er preget av sin rikdom på epikontinentale hav utsatt for kontinental påvirkning. Stratigrafisk er begynnelsen preget av utviklingen av planktoniske foraminiferer og av utseendet til slekten Hantkenina . Den ender sammen med nesten utryddelsen av kokkolitoforen Reticulofenestra reticulata , for rundt 40,4 ± 0,2 millioner år siden, og skylder navnet sitt til det gamle romerske navnet på byen Paris : Lutetia Parisiorum . [ 7 ]
- Bartonian – Startet for 40,4 ± 0,2 millioner år siden. Karl Mayer-Eymar kalte det og definerte grensene i 1857 fra fossilrike leireholdige sedimenteri Sør - England . Stratigrafisk er begynnelsen preget av nesten utryddelse av coccolithophore Reticulofenestra reticulata . Den endte for 37,2 ± 0,2 millioner år siden, preget av den første opptredenen av coccolithophore Chiasmolithus oamaruensis . Navnet kommer fra byen Barton-on-Sea i England . [ 8 ]
- Priabonian : Startet for 37,2 ± 0,1 millioner år siden. Dette er den siste tiden av eocen, hvor Grande Coupure fant sted , en episode med massive utryddelser og markerte faunaendringer. Stratigrafisk er begynnelsen preget av den første opptredenen av coccolithophore Chiasmolithus oamaruensis . Den endte for 33,9 ± 0,1 millioner år siden, og ble preget av planktoniske foraminiferer og utryddelsen av slekten Hantkenina . Navnet kommer fra byen Priabona i Italia . [ 5 ]
Paleogeografi
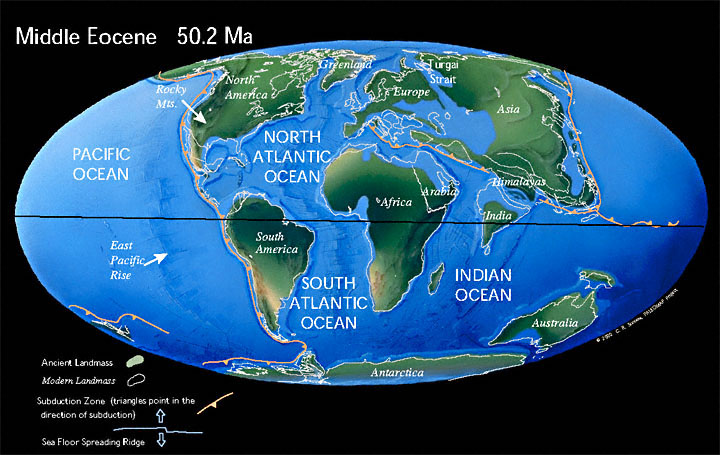
Se også: PangeaDen tredje og siste store fasen av oppløsningen av Pangea - superkontinentet fant sted i tidlig kenozoikum , mellom paleocen og oligocen . Paleokontinentet Laurentia , som består av dagens Nord-Amerika og Grønland , fortsatte å løsrive seg fra Eurasia og utvide det unge Atlanterhavet , til tross for at en viss forbindelse mellom begge landmassene antas fortsatt å ha eksistert. [ 9 ] Ettersom Atlanterhavet fortsatte å ekspandere, fortsatte det gamle Tethyshavet å lukke seg på grunn av tilnærmingen til de afrikanske og eurasiske kontinentene .
I tidlig eocen var det australske kontinentet fortsatt knyttet til Antarktis , men under Lutetian begynte Australia raskt å skille seg fra Antarktis som flyttet nordover på grunn av kontinentaldrift , akkurat som det indiske subkontinentet og New Zealand allerede hadde gjort. Zealand titalls millioner av år tidligere, under kritttiden . Isoleringen av det antarktiske kontinentet ville ha drastiske konsekvenser for det globale klimaet, for eksempel det termiske maksimumet av Paleocen-Eocen eller Azolla-hendelsen .
Orogenese
Den kenozoiske epoken var en periode med intens orogen aktivitet . Under den såkalte alpine orogenien ble fjellene i Tethys-systemet dannet , en rekkevidde som strekker seg over den sørlige delen av Eurasia og inkluderer Alpene , Karpatene , fjellene i Lilleasia , Iran , Hindu Kush , Himalaya , og fjellene i Sørøst-Asia . Denne orogene aktiviteten forårsaket en intens regional metamorfose .
Det indiske subkontinentet, som tidligere hadde brutt seg bort fra Gondwana i den sene krittiden , og siden den gang hadde beveget seg med en hastighet på 16 cm /år, kolliderte med Eurasia i tidlig eocen. Kollisjonen mellom disse to landmassene skapte den høyeste fjellkjeden i verden, Himalaya . [ 10 ] Denne prosessen med orogenese pågår fortsatt i dag, og gjør Himalaya omtrent to centimeter høyere hvert år. [ 11 ]
På den annen side fortsatte den kimmerske orogenien , en orogen prosess som hadde begynt i jura , å skape noen av fjellkjedene som nå finnes i sentrum av det asiatiske kontinentet . Eocen var åstedet for den siste fasen av denne orogenesen.
Eurasia var ikke det eneste kontinentet med orogen aktivitet. Den geologiske konfigurasjonen til mange fjell i Nord-Amerika stammer fra tidlig kenozoikum, for eksempel Black Hills i South Dakota , Wyoming eller Appalacherne på østkysten.
Klima
Det globale klimaet i eocen var sannsynligvis det mest homogene i kenozoikum ; den termiske gradienten fra ekvator til polene var da halvparten av hva den er i dag, og dype havstrømmer var usedvanlig varme. Polarområdene var mye varmere enn i dag, med temperaturer som ligner på dagens nordvestlige USA . De tempererte skogene nådde helt til polene, mens det regnfulle tropiske klimaet nådde opp til 45° nordlig bredde . Den største forskjellen ble funnet i de tempererte breddegrader, selv om klimaet i tropene var likt det i vår tid. [ 12 ] Da det australske kontinentet og Antarktis ble forent til en enkelt landmasse ved begynnelsen av eocen , blandet kalde og varme havstrømmer seg, og opprettholdt en homogen havtemperatur. [ 13 ]
Fra begynnelsen av denne epoken økte temperaturen, i en av de raskeste (i geologiske termer) og ekstreme globale oppvarmingen som noen gang er registrert i geologisk historie, kalt Paleocene – Eocene termiske maksimum . Det var en rask og intens oppvarmingsepisode (opptil 7 °C på høye breddegrader) som varte i mindre enn 100 000 år. [ 14 ] Det termiske maksimumet forårsaket en masseutryddelse, så eocen- og paleocenfaunaene er svært forskjellige.
Det er mulig at denne intense oppvarmingen ble forårsaket av utstøtingen av metanklatrater begravd på havbunnen. Det antas at sedimentene som disse klatratene ble lagret i ble forstyrret etter hvert som havene ble varmet opp, og metanet de inneholdt førte til at den globale temperaturen steg. Det er anslått at to tusen gigatonn metan, en klimagass som er ti ganger kraftigere enn karbondioksid, ble sluppet ut i atmosfæren . [ 15 ]
Økningen i temperaturer over hele planeten var ikke den eneste konsekvensen, ettersom det globale klimaet også ble fuktigere , og mye av denne fuktigheten ble ført til polene. [ 16 ] Den store mengden nedbør over Polhavet , kombinert med konfigurasjonen av kontinentene (som isolerte det fra resten av havene), reduserte saltinnholdet drastisk . Ferskvannet akkumulert i den arktiske sonen forberedte bakken for en stor klimaendring av totalt motsatt fortegn, og det ville markere slutten på eocen.
Klimaet forble varmt under resten av eocen, til tross for at en global avkjøling, som til slutt ville føre til pleistocene isbreer, begynte midt i denne epoken på grunn av to faktorer: Azolla-hendelsen og isolasjonen av Antarktis . .
Azolla-hendelsen fant sted for 49 millioner år siden, da ferskvanns - Azolla - bregnen , også kalt "myggbregne", begynte å vokse i store mengder over Polhavet. Da de sank til havbunnen, ble plantene en del av havbunnssedimentene, hvor de ikke kunne brytes ned på grunn av det lave nivået av oksygen i dypvannslagene. Reduksjonen i mengden karbon i jordens atmosfære bidro til å forvandle planeten fra en "Hothouse Earth" - varm nok til at skilpadder og palmetrær kan bo i polene - til en "Icy Earth". [ 17 ] [ 18 ]
Med separasjonen av det australske kontinentet for rundt førtifem millioner år siden, ble Antarktis fratatt strømmen av ekvatorialvann som inntil da hadde myknet opp klimaet. Med berøvelsen av dette varme vannet, avkjølte Antarktis og Sørishavet begynte å fryse, og skapte en strøm av kaldt vann og isdekker som forsterket den kjølende effekten.
Dyreliv
Fugler
For første gang i jordens historie dominerte fugler over alle andre vesener. Kjempe rovfugler, som Gastornis , tidligere kjent som Diatryma , matet på pattedyr som Propalaeotherium , i Europa og Nord-Amerika , [ 19 ] mens Phorusrhacidae , kjent som "terrorfugler", de ville bli de typiske apex-rovdyrene i Sør-Amerika .
Pingviner , som hadde dukket opp under paleocen , ankom Sør-Amerika rundt midt-eocen , og ved Bartonian hadde de allerede begynt å spre seg over Atlanterhavet. Noen primitive slekter og arter av pingviner er Perudyptes devriesi (76 cm høy) og den store Icadyptes salasi (150 cm høy), begge funnet på ørkenkysten av Ica , i det sørlige Peru ; [ 20 ] likeså Archaeospheniscus eller den gigantiske Anthropornis nordenskjoeldi , som var 170 cm høy og veide rundt 90 kg. Til sammenligning er keiserpingvinen , som er den største levende pingvinen, bare 122 cm høy og 37 kg i vekt .
Anseriformes begynte å diversifisere seg, til slekter som Presbyornis , som var relatert til dagens ender og gjess . Studier av Presbyornithidae- familien er svært viktige for å forstå utviklingen av fugler. Arter som levde i kystområder og var mindre spesialiserte hadde mindre sannsynlighet for å dø ut enn arter som var mer tilpasset et bestemt økosystem .
Palaeotis er en strutsefugl hvis fossiler er funnet på fossilstedet Messel i Tyskland . Disse funnene er spesielt interessante, ettersom den tradisjonelle tesen hevder at strutsefugler har sin opprinnelse i Gondwana , basert på deres nåværende spredning. Oppdagelsen i Europa av Palaeotis , sammen med andre strutsfugler i Nord-Amerika og Mongolia , viser denne hypotesen. [ 21 ]
Bevis for en bredere spredning av papegøyer under eocen er funnet i Quercy - fosforittene . Fossile rester av slekten Quercypsitta , datert mellom 34 og 37 millioner år gamle, indikerer at papegøyer en gang nådde nordligere breddegrader enn de er i dag. [ 22 ] Det eldste papegøyefossilet som er registrert er funnet i Pelsformasjonen , Danmark , og er 54 millioner år gammelt, sammenfallende med begynnelsen av eocen. [ 23 ]
Pattedyr
Den viktigste hendelsen i utviklingen av pattedyr under eocen var trolig utviklingen av hvaler . Etter at deres forfedre forlot livet i vann for 300 millioner år siden, klarte en gruppe pattedyr relatert til de primitive artiodactylene å gjøre overgangen fra et terrestrisk til et vannmiljø.
Denne prosessen begynte med midtre og nedre eocene Pakicetidae i Pakistan . De var kjøttetende landdyr , men konfigurasjonen av ørenes bein og tannsett viste at de representerer det første trinnet i utviklingen av hvaler . Noen få millioner år senere hadde noen skapninger, som Ambulocetus , allerede en amfibisk livsstil , bakbena deres mer tilpasset for svømming enn for å gå på tørt land. [ 24 ] Protocetidae representerer et senere trinn i utviklingen av hvaler, og det er mulig at de da allerede hadde en halefinne som den til moderne hvaler . [ 25 ]
De første fullstendig marine hvaler dukket opp for rundt førtifem millioner år siden. Basilosauridae , som inkluderer slekter som Basilosaurus eller Dorudon , hadde en anatomi veldig lik den til levende hvaler. Likevel var hjernen deres mindre utviklet og de hadde ikke den typiske melonen til odontocetene . De første tannhvalene ville ikke dukke opp før nesten slutten av eocen.
-
Prorastomus ( Sirenia )
-
Ambulocetus ( Cetacea )
-
Basilosaurus ( Cetacea )
Hovdyr fortsatte også å utvikle seg under eocen. Artiodactyls dukket opp i begynnelsen av denne epoken, for femtifire millioner år siden, og ved slutten av eocen hadde de allerede diversifisert seg til de tre gjeldende underordene: Tylopoda ( kameler ), Suinae ( griser ) og Ruminantia ( sauer , geiter , og storfe ) . Den store utvidelsen av Perissodactyla , som fortrengte dem mot mindre velstående habitater , og utseendet til gress i eocen, betinget utviklingen av det spesielle fordøyelsesapparatet som artiodactylene besitter, og som senere ville hjelpe dem med å fjerne Perissodactyla som de dominerende planteetere . . [ 26 ]
De dominerende rovdyrene under eocen var kreodontene . Kjøttetere har vanligvis to kjøttetenner , en jeksel og en premolar , men i stedet var kjøtttennene til kreodonter begge jekslene. [ 27 ] Kreodonter inkluderer noen av de største landlevende pattedyrrovdyrene som noen gang har eksistert, som Andrewsarchus , som vokste til å bli 3,5 meter lang, nesten 2 meter høy og veie 250 kg. [ 19 ] Likevel var deres store størrelse ikke nok til å seire på lang sikt, ettersom de ble overgått av andre rovdyr og endte opp med å bli utryddet i miocen . Deres utryddelse skyldtes en rekke faktorer, for det første var deres lumbosakrale ledd ikke tilstrekkelig utviklet til å løpe som andre rovdyr, og dessuten gjorde deres plantigrade tilstand dem mindre effektive når det kom til løping. [ 28 ] For det andre hadde kreodonter forskjellig tannsett som gjorde dem til obligatoriske rovdyr , det vil si at de var begrenset til kun å spise kjøtt, mens miaciner og de fleste rovdyr på den tiden fortsatt hadde tenner tilpasset til å tygge andre typer mat. [ 27 ] Den siste kreodonten som dukket opp i fossilregistret , Dissopsalis , ble utryddet for åtte millioner år siden.
Fisk
Under eocen gjennomgikk lamniforme haier (eller stripete haier) en stor diversifisering. Goblinhaien er et av mange eksempler på haier som dukket opp i løpet av denne tiden . [ 29 ] En av de mest bemerkelsesverdige artene var Otodus obliquus , en hai som dukket opp i paleocen , som kunne bli ni meter lang og livnærte seg av sjøpattedyr, fisk og andre haier. [ 30 ] Mange paleontologer mener at O. obliquus er en stamfar til slekten Charcharocles , og derfor nært beslektet med den største rovhaien som noen gang har levd, Carcharodon megalodon . Andre relaterer imidlertid O. obliquus til den store hvithaien , selv om antallet paleontologer som tror den er avtagende, ettersom det er flere og flere bevis på dens forhold til megalodonen. [ 31 ]
En annen viktig fisk var Enchodus , et rovdyr relatert til laks . Enchodus hadde en serie støttenner foran på over- og underkjeven og på palatinebeina . Til tross for at det er et rovdyr, er det store flertallet av fossilene funnet inne i magen til andre større rovdyr, for eksempel mosasaurider , plesiosaurer eller sjøfuglen Baptornis .
Reptiler
En av de største kjente slangeartene levde under eocen. Gigantophis garstini kunne ha overskredet ti meter i lengde, mens de største nåværende slangene, anacondas , er rundt syv meter lange. Denne slangen, som bebodde planeten for førti millioner år siden i området i dagens Egypt , matet sannsynligvis av basale proboscideans , forfedrene til dagens elefanter . [ 32 ] En annen stor slange som levde i løpet av denne tiden var Palaeophis , en sjøorm. Opprinnelig ble det anslått at lengden varierte mellom tretti og førti meter, et enormt høyt tall for en slange, men nyere estimater viser lengder på omtrent ni meter. [ 33 ]
Leddyr
Det mest relevante i eocen i forhold til leddyrene var maurenes ekspansjon . Under kritttiden var det bare noen få arter av maur på det gamle kontinentet Laurasia , som representerte mindre enn 1% av alle insekter . På slutten av paleocen begynte en adaptiv stråling som varte under eocen og gjorde dem til de dominerende insektene til slutten av epoken. Suksessen hans var slik at 90 % av maurartene som levde under eocen fortsatt overlever. [ 34 ]
Fossiler av Formicium , en slekt av maur som omfatter totalt fem arter og som bare er kjent fra restene av hanner og dronninger , er funnet på fossilområdet Messel i Tyskland . Vingespennet til dronningene varierte fra 13 til 15 centimeter, noe som gjør den til den største mauren som noen gang har levd. Fossiler av denne slekten er også funnet i Storbritannia og USA , men i disse tilfellene er det kun isolerte vinger .
Flora
I tidlig eocen varmet høye temperaturer havene og skapte et varmt, fuktig miljø, der skoger kan bli funnet som strekker seg fra pol til pol. Bortsett fra de tørreste og mest ekstreme ørkenområdene , var jorden fullstendig dekket av skog. [ 13 ]
Polarskogene fikk en flott utvidelse. Fossiler og til og med bevarte rester av trær , som Cupressaceae eller Metasequoia - slekten , er funnet på Ellesmere Island , som ligger i det kanadiske arktiske området. De bevarte restene som er funnet er ikke fossiler, men originale fragmenter som ble bevart i oksygenfattige vann i eocene sumpskoger og senere begravd før nedbrytningen startet. [ 35 ] Fossiler av subtropiske og til og med tropiske eocene trær er også funnet på steder som Grønland eller Alaska . Jungle nådde så langt nord som det nordvestlige USA og Europa . [ 13 ]
I begynnelsen av denne tiden vokste det palmer i Alaska og Nord-Europa, men etter hvert som tiden gikk, og planeten ble avkjølt, ble det mindre rikelig med palmer. Metasequoia var vidt distribuert.
Avkjølingen begynte midt i sesongen. Ved slutten av eocen hadde det indre av kontinentene allerede begynt å tørke ut, og i enkelte områder begynte skogene å krympe betraktelig. Gresset , som nettopp hadde dukket opp, var begrenset til bredden av elvene og hadde ennå ikke spredt seg over savannene og slettene . [ 36 ]
Terrestrisk avkjøling ble ledsaget av sesongmessige endringer . Løvtrær , som var mer tilpasset store endringer i temperaturen, begynte å råde over tropiske eviggrønne arter. Ved slutten av eocen dekket løvskoger allerede store områder på de nordlige kontinentene, inkludert Nord-Amerika , Eurasia og Arktis , mens jungelen bare klarte å motstå i Sør-Amerika , India og Australia .
Antarktis , som startet eocen innhyllet i tempererte-subtropiske skoger, avkjølte seg betydelig ettersom tiden gikk. Den tropiske floraen med høy temperatur forsvant, og ved tidlig oligocen var det antarktiske kontinentet allerede hjemsted for løvskog og store områder med tundra .
Stehlins store pause
Det store Stehlin-bruddet (på fransk Grande Coupure) var en utryddelseshendelse som førte til en stor endring i bestanden av forskjellige organismer i Europa , med pattedyr som en av de mest berørte gruppene. Den sveitsiske paleontologen Hans Georg Stehlin skapte navnet i 1910 , [ 37 ] med henvisning til den drastiske endringen i europeiske pattedyr.
Denne hendelsen, som fant sted for 33,9 millioner år siden, [ 38 ] har fungert som et kriterium for å definere grensen mellom eocen og oligocen , og er preget av store utryddelser og allopatrisk arter av isolerte primitive arter. En lignende begivenhet fant sted i Asia, som ble kalt "Mongolian Remodeling".
En av hovedårsakene til dette faktum ser ut til å være stengingen av Turgai-stredet , som forente Europa og Asia og satte en stopper for den paleogeografiske isolasjonen av Europa, og dermed tillot massive migrasjoner av arter mellom de to kontinentene. I tillegg fremhevet åpningen av Drake-passasjen den antarktiske sirkumpolare strømmen , og initierte dermed progressiv avkjøling, noe som førte til dannelsen av en iskappe i Antarktis, [ 39 ] samt dannelsen av et lag med kaldt vann på havbunnen . Dannelsen av hetter forårsaket et betydelig fall i havnivået og fremhevet albedoeffekten , som reflekterte solstråling og forårsaket et stort fall i temperaturer. Laget med kaldt vann førte til at mange av artene som bodde i varmt vann (som Basilosaurus og noen haier) gikk til grunne, noe som ga opphav til en svært dårlig diversifisert fauna. Klimaendringene som fant sted ville være innledningen til de første polare istidene .
Pattedyr gjennomgikk en stor renovering. Taxaene til endemiske europeiske pattedyr ble erstattet av asiatiske immigranter , og gjorde dermed mer enn halvparten av europeiske pattedyr utryddet, og dermed påvirket resten av floraen og faunaen. Bløtdyr gjennomgikk også en større renovering, spesielt på vestkysten av USA og i kystsletten i den nordlige Mexicogulfen . [ 40 ]
Meteorittnedslag
Noen teorier peker på virkningen av ildkuler over Sibir som hovedårsaken til denne hendelsen på grunn av anomalier funnet i spor av iridium , et veldig nyttig element for å oppdage meteorittnedslag , angivelig for 34 millioner år siden. [ 41 ] [ 42 ] Det ble opprinnelig foreslått som en enkelt hendelse, men senere ble det foreslått et forskjøvet masseutryddelsesmønster, som hver var forskjellige kometer eller meteornedslag . [ 43 ] [ 44 ] [ 45 ] Denne forskjøvede ekstinksjonsmodellen ble imidlertid senere forkastet da det ble vist at påvirkningene var før utryddelsene, [ 46 ] senere bekreftet takket være studiet av en rekke kutt og undersøkelser oseanisk. [ 47 ] Disse sonderingene er svært nøyaktig datert av planktoniske foraminiferer , og konkluderer med at deres utryddelse ikke var forårsaket av nedslag. [ 47 ] [ 48 ]
Den påfølgende oppdagelsen av kvarts med sjokkmetamorfosmer og nikkelrike spineller , [ 49 ] [ 50 ] [ 51 ] samt oppdagelsen av flere kratere i Sibir ( Popigai-krateret ) og Nord-Amerika , har bekreftet at det for omtrent 35 år siden , 6 millioner år siden tre store meteoritter traff planeten. [ 52 ] Noen forfattere har antydet at disse påvirkningene akselererte global avkjøling, [ 53 ] [ 54 ] selv om isotopdata ikke støtter denne akselerasjonen. [ 39 ]
Paleontologiske steder
Dette er noen av de mest fremragende paleontologiske stedene fra eocen-tiden:
Wadi Al-Hitan
Wadi Al-Hitan ( arabisk وادي الحيتان, hvalenes dal ) er en region i den vestlige ørkenen i Egypt , hvor viktige fossiler av tidlige hvaler er funnet . Disse restene representerer en av hovedregistrene i historien om utviklingen av arter: overgangen til hvaler fra landdyr til vannlevende dyr . Fossilene som er funnet i Wadi al-Hitan lar oss vite at under eocen var det som nå er Sahara-ørkenen da et grunt hav av Tethyshavet .
Komplette eller nesten komplette Zeuglodon- fossiler er de mest fremtredende funnene i dalen, og gir den dermed kallenavnet "Zeuglodondalen". Dorudon er en annen forhistorisk hval som det er funnet fossiler for i Wadi al-Hitan . I 2005 ble dalen erklært som et verdensarvsted av UNESCO .
Green River Formation
Green River Formation er en geologisk formasjon som strekker seg over nordlige Utah , vestlige Colorado og sørvestlige Wyoming , i USA . I den kan du finne to godt differensierte områder med veldig fin siltig gjørme , der et stort mangfold av komplette og detaljerte fossiler er bevart. Den mest produktive sonen, den såkalte "18-tommerssonen", inneholder et stort antall fossiler, spesielt fisk , i en serie laminerte lag av gjørme, og representerer omtrent fire tusen år med avsetninger. Den andre fossilholdige sonen er en ulaminert sone som er litt mindre enn to meter tykk, hvor det også kan finnes svært detaljerte fossiler, selv om utvinningen er vanskelig fordi den ikke er ordnet i separerbare laminater. [ 55 ]
Marambio Island
Marambio Island, også kalt Seymour Island, er en av seksten store øyer som omgir spissen av den antarktiske halvøya .
Den norske kapteinen Carl Anton Larsen oppdaget flere fossiler på øya i 1882 , på sin første tur til Marambio-øya, ombord på skipet Jason . Siden den gang har øya vært gjenstand for en rekke paleontologiske undersøkelser.
Forskning utført på øya Marambio har bidratt til å forstå avkjøling under eocen, en prosess som kulminerte i starten av istiden i Antarktis . Studier av karbonat fra forskjellige deler av Sørishavet tyder på at det, snarere enn en monoton avkjøling i temperaturen i denne epoken, var en kort oppvarmingsepisode i midten av eocen. Mange fossiler har også blitt studert, for eksempel noen utdødde pingviner , forskjellige muslinger og andre dyr og planter . [ 56 ]
London Clay
London Clay er en marin forekomst kjent for sine fossiler. Det er det viktigste fossilrike stedet for Ypresian i Sør - England , og det er det eneste stedet i Europa hvor et enormt utvalg av plantefossiler fra Nedre Eocen kan bli funnet.
Leiren ble avsatt i et hav som var to hundre meter dypt i det østligste området. Opptil fem sykluser med avsetning (som representerer en overskridelse etterfulgt av en reduksjon i havdybden) er funnet, for det meste i den grunnere vestlige regionen. Hver syklus begynner med grovt materiale (inkludert avrundede flintkonsentrasjoner ), og slutter med stadig mer sandholdig leire. [ 57 ]
Dyrefossiler som er funnet inkluderer muslinger , gastropoder , nautilus , krabber , hummer, fisk (inkludert haitenner ), krypdyr ( spesielt skilpadder ) og sjeldne fugleeksemplarer . Noen pattedyrfossiler er også funnet .
Plantefossiler, inkludert frukt og frø , er svært rikelig i London Clay . Plantefossiler begynte å bli samlet for litt mindre enn tre århundrer siden, og rundt 350 arter er blitt beskrevet . Dette gjør floraen i London Clay til en av de mest varierte når det gjelder frukt og fossile frø. [ 58 ]
Messel fossilside
Messel Fossil Site, som ligger i Hessen , Tyskland , og erklært et verdensarvsted i 1995 , er et tidligere oljeskiferbrudd som inneholder den best bevarte prøven av mellomeocene fauna og flora kjent. På de fleste steder er det å finne delvise skjeletter en prestasjon, men i Messel er det mange tilfeller av omfattende bevaring, og noen bevarer til og med pelsen , fjærene eller hudmerkene til noen arter. Det store artsmangfoldet er også et pluss, blant annet takket være gassutbrudd. I innskuddet er det funnet:
- Mer enn ti tusen fossilisert fisk som tilhører forskjellige arter.
- Tusenvis av både vannlevende og terrestriske insekter , hvorav noen av fargene er bevart.
- Store mengder små pattedyr , inkludert pygmehester , store gnagere , primater , opossum , beltedyr , slektninger av jordvarken og flaggermus .
- Tallrike fugler , spesielt rovdyr.
- Krokodiller , frosker , skilpadder , salamandere og andre krypdyr og amfibier .
- Mer enn tretti planterester , som palmeblader , frukt , pollen og nøtter .
Se også
Notater og referanser
- ↑ Fargene tilsvarer RGB-kodene godkjent av International Commission on Stratigraphy . Tilgjengelig på nettstedet til International Commission on Stratigraphy, i «Standard Color Codes for the Geological Time Scale» .
- ↑ Global Boundary Stratotype Section and Point (GSSP) av International Commission of Stratigraphy , status 2009.
- ↑ Internasjonalt stratigrafisk diagram, 2008
- ^ a b Rohde, Robert A. (2005). "Ypresian ICS Stage" . GeoWhen Database . Den internasjonale kommisjonen for stratigrafi. Arkivert fra originalen 28. november 2015 . Hentet 10. mai 2008 .
- ^ a b Rohde, Robert A. (2005). "Priabonian ICS Stage" . GeoWhen Database . Den internasjonale kommisjonen for stratigrafi. Arkivert fra originalen 28. november 2015 . Hentet 10. mai 2008 .
- ↑ a b Tradisjonelt har tertiær og kvartær blitt brukt i stedet for den nåværende kenozoikum , med en rekke erathems eller epoker, med bruk av kenozoikum som et synonym for tertiær og delt i sin tur inn i paleogen og neogen. Tertiær og kvartær kan også bli funnet som underaldre innenfor erathem eller kenozoikum. For øyeblikket, i 2009, har begrepet tertiær (og den tilsvarende underavdelingen innen kenozoikum) sluttet å bli anbefalt av Den internasjonale kommisjonen for stratigrafi for global skala, og etterlater kenozoikum delt inn i paleogene, neogene og kvartære systemer eller perioder.
- ^ Rohde, Robert A. (2005). "Lutetian ICS Stage" . GeoWhen Database . Den internasjonale kommisjonen for stratigrafi. Arkivert fra originalen 24. mai 2008 . Hentet 10. mai 2008 .
- ^ Rohde, Robert A. (2005). "Bartonian ICS Stage" . GeoWhen Database . Den internasjonale kommisjonen for stratigrafi. Arkivert fra originalen 4. februar 2008 . Hentet 10. mai 2008 .
- ^ "Eocenens tektonikk" . UC Museum of Paleontology (på engelsk) . 1999 . Hentet 18. mai 2008 .
- ^ Bin Zhu, William SF Kidd, David B. Rowley, Brian S. Currie, Naseer Shafique (2006). "Alder for initiering av India-Asia-kollisjonen i det øst-sentrale Himalaya" (PDF) . Journal of Geology 114 . s. 641-643 . Arkivert fra originalen 2. desember 2005.
- ^ Jean-Louis Mugnier, Pascal Leturmy, Gérard Vidal (2004). "Kinetikk og sedimentær balanse i Subhimalayan-sonen, Vest-Nepal" . Skyvetektonikk og hydrokarbonsystemer 82 . s. 115-130 .
- ^ Stanley, Steven M. (1999). Earth System History . New York: W. H. Freeman and Company. ISBN 0-7167-2882-6 .
- ↑ abcAgaric ( 2007 ). "Jordens eocene epoke" . Tilknyttet innhold . Arkivert fra originalen 2. januar 2013 . Hentet 10. mai 2008 .
- ↑ Gavin A. Schmidt, Drew T. Shindell (2003). "Atmosfærisk sammensetning, strålingspådriv og klimaendringer som en konsekvens av en massiv metanfrigjøring fra gasshydrater" . Paleoceanografi 18 (1). s. 1004, http://dx.doi.org/10.1029/2002PA000757 . Arkivert fra originalen 20. oktober 2011 . Hentet 10. mai 2008 .
- ↑ White, Kasey (2003). Ledetråder til global oppvarming . UC Santa Cruz . Arkivert fra originalen 6. juli 2008 . Hentet 10. mai 2008 .
- ↑ Pagani, M.;Pedentchouk, N.; Huber, M.; Sluijs, A.; Schouten, S.; Brinkhuis, H.; Sinninghe Damste, JS; Dickens, G.R.; Andre (2006). "Arktisk hydrologi under global oppvarming ved Palaeocene/Eocene termiske maksimum" . Nature 443 (7111). s. 598 .
- ^ Macdougall, Doug (2004). Frozen Earth: The Once and Future Story of Ice Ages . University of California Press . ISBN 0-520-24824-4 .
- ↑ Mulvaney, Kieran (2001). At the Ends of the Earth: A History of the Polar Regions . Washington, DC: Island Press. ISBN 1-55963-908-3 .
- ^ a b Haines, Tim (2001). Vandre med dyr . BBC Books. ISBN 0-563-53763-9 .
- ^ "Forhistoriske kjempepingviner" . Verden er. 26. juni 2007 . Hentet 5. oktober 2008 .
- ↑ Chatterjee, Sankar (1997). The Rise of Birds . Johns Hopkins University Press . ISBN 0-8018-5615-9 .
- ^ Mourier-Chauviré, Cécile (1992). "Une nouvelle famille de Perroquets (fugler, Psittaciformes) dans l'Eocène supérieur des Phosphorites du Quercy". Geobios, Memoire Special 14 . s. 169-177 .
- ↑ David M.Waterhouse, Bent E.Lindow, Nikita V.Zelenkov, Gareth J.Dyke (2008). "To nye papegøyer (Psittaciformes) fra den nedre eocene pelsformasjonen i Danmark" . Paleontologi 51 (3). s. 575-582 .
- ↑ Sutera, Raymond (2002). "Hvalens opprinnelse og kraften til uavhengige bevis" . Rapporter fra Nasjonalt senter for realfagsutdanning 20 (5). s. 33-41 .
- ^ Clementz, Marc T., Goswami, A.; Gingerich, P.; Koch, P. (2006). Isotopiske registreringer fra tidlige hvaler og sjøkyr: kontrasterende mønstre av økologisk overgang. Journal of Vertebrate Paleontology 26 (2). s. 355-370 .
- ^ Janis, Christine; Jarman, Peter (1984). The Encyclopedia of Pattedyr . New York: Fakta på filen. ISBN 0-87196-871-1 .
- ^ a b MacDonald, David (1993). "The Carnassial Connection" . Velvet Claw: A Natural History of the Carnivores . BBC Books. ISBN 0-563-20844-9 .
- ↑ William Harmon Norton. Kapittel XXI. Tertiæren» (eBok) . The Elements of Geology (på engelsk) . Globez Publishing. Arkivert fra originalen 9. mars 2008 . Hentet 20. mai 2008 .
- ^ Sepkoski, Jack (2002). "Et kompendium av fossile marine dyreslekter (Chondrichthyes-oppføring)" . Bulletins of American Paleontology 364 . s. 560 . Arkivert fra originalen 10. mai 2012.
- ^ Renz, Mark (2002). Megalodon: Hunting the Hunter . s. 26-30, Paleo Press. ISBN 0-9719477-0-8 .
- ↑ David Ward, Jim Bourdon (2007). "Carcharocles" (på engelsk) . Elasmo.com . Hentet 20. mai 2008 .
- ^ New Scientist (2004). «En gigant blant slanger» . New Scientist (2473). s. 17 .
- ↑ Dennis Parmley, Melanie DeVore (2005). "Palaeopheid-slanger fra den sene eocene Hardie Mine Local Fauna of Central Georgia" . Sørøstnaturforsker 4 (4). s. 703–722 .
- ↑ David Grimaldi, Donat Agosti (2000). "En formisin i New Jersey kritt rav (Hymenoptera: Formicidae) og tidlig evolusjon av maurene" . Proceedings of the National Academy of Sciences 97 (25). s. 13678–13683 .
- ↑ "Eocene Dawn Redwood Fossil Frond fra Cache Creek" . Det virtuelle fossilmuseet . Hentet 20. mai 2008 .
- ↑ Thomas R. Van Devender, Mark A. Dimmitt. "Ørkengress" . Arizona-Sonora Desert Museum . Hentet 20. mai 2008 .
- ^ H. G. Stehlin (1910). "Remarques sur les faunules de Mammifères des couches eocenes et oligocenes du Bassin de Paris". Bulletin of the Société Géologique de France 4 (9). s. 488-520 .
- ↑ Lutherbacher, HP; Ali, JR; Brinkhuis, H.; Gradstein, FM; Hooker, JJ; Monechi, S.; Ogg, JG; Powell, J.; Rolle, U.; Sanfi lippo, A; Schmitz, B. (2004). "Den paleogene perioden". En geologisk tidsskala 2004 . Cambridge University Press , Cambridge, s. 384-408. ISBN 0-521-78142-6 .
- ↑ a b Livermore, Roy; Nankivell, Adrian; Eagles, Graeme; Morris, Peter (2005). "Paleogen åpning av Drake Passage" . Earth and Planetary Science Letters 236 (1-2). s. 459-470 .
- ^ Ivany LC; Patterson WP; Lohmann KC (2000). "Kjøligere vintre som en mulig årsak til masseutryddelser ved grensen mellom eocen og oligocen" . Natur 407 . s. 887-890 .
- ^ Ganapathy, R. (1982). "En stor meteorittpåvirkning på jorden for 65 millioner år siden: Bevis fra kritt-tertiær grenseleire" . Science 216 (4459). s. 885-886 .
- ↑ Álvarez, W., Asaro, F., Michel, HV, Álvarez, LW (1982). "Iridium-anomali tilnærmet synkron med terminale Eocene-utryddelser" . Science 216 (4548). s. 886-888 .
- ↑ Hut, P.; Alvarez, W.; Eldste, W.P.; Hansen, T.; Kauffman, EG; Keller, G.; Skomaker, EM; Weissman, P.R. (1987). "Kometdusjer som en årsak til masseutryddelser" . Natur 329 . s. 118-126 .
- ↑ Keller, G. (1986). "Trinnvise masseutryddelser og påvirkningshendelser: Sen eocen og tidlig oligocen". Marin mikropaleontologi 13 . s. 267-293 .
- ^ Kauffman, E.G. (1988). "Dynamikken til marin trinnvis masseutryddelse". Spansk tidsskrift for paleontologi (ekstraordinært). s. 57-71 .
- ↑ Keller, G.; D'Hondt, SL; Orth, CJ; Gilmore, JS; Oliver, PQ; Skomaker, EM; Molina, E. (1987). "Sen eocen-påvirkningsmikrosfærer - Stratigrafi, alder og geokjemi" . Meteoritikk 22 (mars). s. 25-60 .
- ↑ a b Molina, E.; Gonzalvo, C.; Keller, G. (1993). "Den eocen-oligosen planktiske foraminiferale overgangen: utryddelser, påvirkninger og hiatuser" . Geologisk magasin 130 (4). s. 483-499 .
- ^ Gonzalvo, C. og Molina, E. (1992). "Biostratigrafi og kronostratigrafi av Eocene-Oligocene transitt i Torre Cardela og Massignano (Italia)". Spanish Journal of Paleontology 7 (2). s. 109-126 .
- ↑ Clymer, Aron K.; Bice, David M.; Montanari, Alessandro (1996). "Sjokkert kvarts fra slutten av eocen: Slagbevis fra Massignano, Italia" . Geologi 24 (6). s. 483-486 .
- ↑ Pierrard, O.; Robin, E.; Rocchia, R.; Montanari, A. (1998). "Utenomjordisk Ni-rik spinell i øvre eocene sedimenter fra Massignano, Italia" . Geologi 26 (4). s. 307-310 .
- ↑ Molina, E.; Gonzalvo, C.; Ortiz, S.; Cross, L.E. (2006). Foraminiferal omsetning over Eocen-Oligocene-overgangen ved Fuente Caldera, Sør-Spania: ingen årsak-virkningsforhold mellom meteorittnedslag og utryddelser. Marin mikropaleontologi 58 . s. 270-286 .
- ↑ Poag, C.W.; Mankinen, E.; Norris, R.D. (2003). "Sene eocenpåvirkninger: geologiske rekorder, korrelasjon og paleomiljømessige konsekvenser". Fra drivhus til ishus . Columbia University Press , New York, s. 495-510. ISBN 0-231-12716-2 .
- ↑ Wonhof, HB; Smit, J.; Brinkhuis, H.; Montanari, A.; Nederbragt, A.J. (2000). "Global avkjøling akselerert av tidlig sen eocen-påvirkning" . Geologi 28 (8). s. 687-690 .
- ↑ Bodiselitsch, B; Montanari, A.; Koeberl, C.; Coccioni, R. (2004). "Forsinket klimaavkjøling i sen eocen forårsaket av flere påvirkninger: høyoppløselige geokjemiske studier ved Massignano" . Earth and Planetary Science Letters 223 (3-4). s. 283-302 .
- ↑ Kaytee Brenes, Micah Fleming, Nalini Rao, Lani Smith (1999). "The Green River Formation" . Lokaliteter i eocen . Hentet 21. mai 2008 .
- ↑ Ivany, LC; Lohmann KC; Hasiuk F. (2005). "Oppvarming av middels eocen på Seymour Island, Antarktis: Paleotemperaturer på kontinentalsokkelen registrert i bløtdyrkarbonater" . Årsmøte i Salt Lake City (oktober). s. 16-19 . Arkivert fra originalen 21. august 2008 . Hentet 21. mai 2008 .
- ^ Sumbler MG (1996). London og Thames Valley . British Regional Geology-serien, British Geological Survey. ISBN 0-11-884522-5 .
- ^ Collinson, M. (1983). Fossile planter av London Clay . Den paleontologiske foreningen. ISBN 0-901702-26-9 .
Bibliografi
På spansk
- Lopez Martinez , Nieves (1998). Geologi og paleontologi av eocen av La Pobla de Segur, Lleida . Universitetet i Lleida. ISBN 84-89727-60-0 .
På engelsk
- Donald R. Prothero (1993). The Eocene-Oligocene Transition: Paradise Lost . Columbia University Press. ISBN 0-231-08091-3 .
- Donald R. Prothero, Robert J. Emry (1996). Den terrestriske eocen-oligocene-overgangen i Nord-Amerika . Cambridge University Press. ISBN 0-521-43387-8 .
- Marie-Pierre Aubry, William A. Berggren, Marie-Pierre Aubry, Spencer G. Lucas (1998). Sent paleocen-tidlig eocen biotiske og klimatiske hendelser i marine og terrestriske registreringer . Columbia University Press. ISBN 0-231-10238-0 .
- Gregg F. Gunnell (2001). Eocen biologisk mangfold: uvanlige forekomster og sjeldne utvalgte habitater . Kluwer Academic Pub. ISBN 0-306-46528-0 .
- Elizabeth Nesbitt, Donald R. Prothero, Linda C. Ivany (2003). Fra drivhus til ishus: The Marine Eocene-Oligocene Transition . Columbia University Press. ISBN 0-231-12716-2 .
På tysk
- Von Koenigswald, W.; Storch, G.; Habersetzer, J. (1997). Messel. Ein Pompeji der Paläontologie . Jan Thorbecke Verlag, Stuttgart. ISBN 3-7995-9083-8 .
Eksterne lenker
 Wikimedia Commons er vert for en mediekategori for eocen .
Wikimedia Commons er vert for en mediekategori for eocen .- Eocen på Celestia-nettet
- Eocen paleomap
- Livet i eocen
- Eocen og Eocen-Oligocene overgangen
- Wadi Al-Hitan (Hvaldalen) på UNESCOs verdensarvliste (på engelsk)
- Green River- formasjonen
- Messel Fossil Site (på tysk)
{kind=link}